|
|
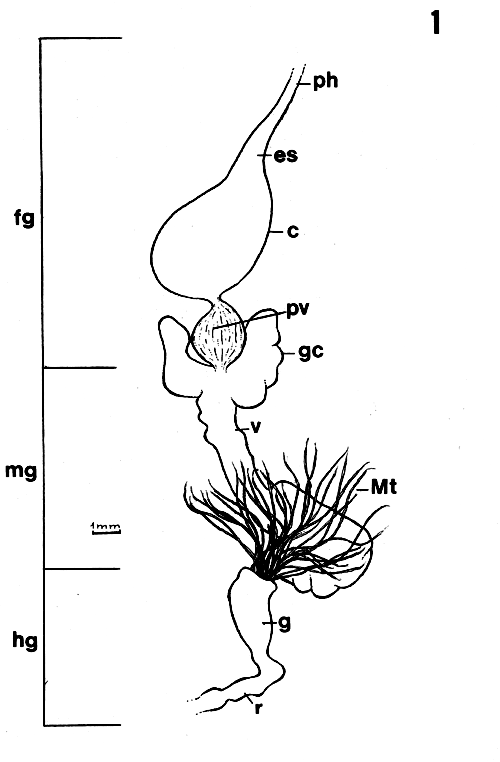
| Figure 1. General morphology of digestive tract of Endecous spp. c = crop; es = esophagus; fg = foregut; g = gut; gc = gastric caecum; hg = hindgut; mg = midgut; Mt = Malpighian tube; ph = pharynx; pv = proventriculus; r = rectum; v = ventriculus. |
|
Entomotropica |
ISSN 1317-5262 |
Carmem S Fontanetti1, Edison Zefa2, Fábio Passetti3, Alejo Mesa1
1Departamento de Biologia, Instituto de Biociências, UNESP. Av. 24-A, no 1515, 13506-900 Rio Claro, SP, Brasil. E-mail: [email protected]Recibido: 08-xi-2000
Aceptado: 19-iii-2001
Correcciones devueltas por el autor: 11-i-2002
Fontanetti CS, Zefa E, Passetti F, Mesa A. 2002. Morphological characterization and comparative analysis of the proventriculus from three species of Endecous Saussure, 1878 (Orthoptera: Gryllidae: Phalangopsinae). Entomotropica 17(1):15-23.
The diversity in the morphology of the digestive tract of insects has led several authors to use it as an auxiliary character in systematics and phylogeny, with special emphasis on the proventriculus, whose morphological variability is related to the diversity of insect feeding habits. In the present study, using scanning electron microscopy, we characterized the proventriculus of six populations of three Endecous species collected in cerrados, woods, fields, forests and caves in order to determine whether this structure suffered some divergence. General morphology was found to be the same for all species analyzed, with the presence of six internal rows of sets of sclerotized appendices formed by a central projection, called middle tooth, and by two lateral expansions called lateral teeth. Both middle and lateral teeth presented apical projections, called denticles, whose number varied from specimen to specimen. Small differences in the shape of the apex of lateral and middle teeth were observed, with the latter being more rounded in cave crickets and more sharp in epigean specimens. Although no studies have been conducted to specifically determine the digestive content of the species analyzed, it is clear that the food available in forests differs from that available in caves both in terms of quality and texture. The authors believe that these differences, although subtle, may reflect an adpatation to variations in diet to which these populations were submitted in epigean and hypogean regions.
Additional key words: Crickets, insect anatomy.
Fontanetti CS, Zefa E, Passetti F, Mesa A. 2002. Caracterización morfológica y análisis comparativo de tres especies de Endecous Saussure, 1878 (Orthoptera: Gryllidae: Phalangopsinae). Entomotropica 17(1):15-23.
La diversidad en la morfología del tracto digestivo de los insectos condujo a varios autores a utilizarlo como carácter adicional en estudios de sistemática y filogenia. El proventrículo, cuya variabilidad morfológica está relacionada a la dieta de los insectos, ha recibido especial énfasis. En el presente trabajo se caracteriza con la ayuda de microscopía electrónica de barrido el proventrículo de seis poblaciones pertenecientes a tres especies de Endecous Saussure, encontradas en ambientes de cerrado, bosque, campo abierto, selva y cuevas, para poder determinar si existen diferencias en el proventrículo. La morfología general observada es la misma en las tres especies e incluye seis hileras internas de series de apéndices esclerotizados formados por una proyección central, llamada «diente medio», y por dos expansiones laterales llamadas «dientes laterales». Tanto los dientes medios como los laterales presentan proyecciones apicales llamadas «dentículos», cuyo número varía individualmente. Se observaron pequeñas diferencias en la forma del ápice de los dientes laterales y medios. Estos últimos son más redondeados en grillos cavernícolas y más afilados en ejemplares epigéicos. No se hicieron estudios para determinar el contenido digestivo de las especies examinadas, pero es obvio que el alimento disponible en bosques difiere de aquel en cuevas, tanto en calidad como en textura. Los autores creen que estas diferencias, aún siendo sutiles, podrían reflejar adaptaciones a variaciones en la dieta a las cuales están sometidas las poblaciones epigéicas e hipogéicas, respectivamente.
Palabras clave adicionales: Anatomía de insectos, grillos.
The diversity in the morphology of the digestive tract of insects has led several authors to use it as an auxiliary character in the systematics and phylogeny of several taxonomic groups (Bordas 1896, 1898; Mortiner 1965; Singh and Judd 1966; Grant and Rentz 1967; Deligne 1971). Special attention has been devoted to the proventriculus, a transition area between the foregut and midgut, because of its morphologic variability usually related to the feeding habits of the insects. The relationship between morphology of the proventriculus and feeding habits has been well established in termites (Lebrun 1985; Lebrun and Lequet 1985), in some Trichoptera (Gibbs 1967) and in a few Orthoptera species (Judd 1948; Bland and Rentz 1991; Fontanetti and Zefa 2000).
The objective of the present study was to describe the morphology of the proventriculus and possible differences in this structure in three species of Endecous: E. itatibensis Rehn, 1918, E. cavernicolus Costa-Lima, 1940 and E. betariensis Melo and Pellegatti-Franco, 1998 from six populations found in different habitats.
Endecous are crickets of the subfamily Phalangopsinae distributed mainly in tropical areas, where they are found in the native vegetation, in leaf litter, under logs and rocks, on rock cliffs and even in abandoned habitations (Otte and Alexander 1983); they have nocturnal habits and show pre-adaptations that allow them to live in caves, changing to a troglophile condition.
In Brazil, these insects are found in cerrados, woods, open fields, semidecidual forest, alluvial semidecidual forest and caves.
The material used was collected by E. Zefa and collaborators and deposited in the collection of the Department of Biology, UNESP - Rio Claro, São Paulo, Brasil.
i) Endecous itatibensis Rehn, 1918 - 2 males and 4 females collected in "cerrado" vegetation in the municipal district of Corumbatai, São Paulo, on May 10, 1996; 8 males and 6 females, collected at Cave of Fazendão: lat 22o 24'37"S, long 47o 47'34"W, on November 25, 1997, municipal district of Ipeúna, São Paulo; 3 males, collected in alluvial semidecidual forest, municipal district of Piracicaba, São Paulo, ESALQ Campus (Escola Superior de Agronomia "Luiz de Queiroz"), on November 3, 1999; 3 males and 1 female, collected in semidecidual forest, municipal district of Itatiba, São Paulo, Km 95, Highway D. Pedro I, on November 20, 1999.
ii) E. cavernicolus Costa-Lima, 1940 - 3 males, collected at Cave of Lapinha: lat 19o 33'40"S, long 43o 57'30"W, municipal district of Lagoa Santa, Minas Gerais, on May 22, 1998;
iii) E. betariensis Mello e Pellegatti-Franco, 1998 - 2 males collected in the municipal district of Iporanga, São Paulo, PETAR (Parque Turístico do Alto do Ribeira), Cave of Alambari de Cima: lat 24o 33'15"S, long 48o 39'50"W; 10 males and 1 female, collected at Cave of Lage Branca: lat 24o32'57"S¨, long 48o 43'15"W, October 4 to 6, 1997.
The digestive tract of adult males and females was dissected in physiological solution. This structure was drawn with the help of a camera lucida coupled to a stereomicroscope. The proventriculus was fixed in Karnowsky (1965) solution and processed for observation by scanning electron microscopy JEOL ISM-P.15 of Department of Biology, UNESP, Rio Claro, SP, Brasil.
1. General morphology of the digestive tract
The digestive tract of the species studied showed the basic composition observed in insects (Figure 1): foregut (fg) consisting of pharynx (ph), esophagus (es), crop (c) and proventriculus (pv), midgut (mg) or ventriculus (v), with a pair of gastric caeca (gc) and hindgut (hg) whose beginning is marked by the insertion of Mapighian tubules (Mt), ending in the rectum (r) and anal area.
When the proventriculus of males and females of each population was compared no differences were detected. The general morphology of the digestive tract as well as that of the proventriculus proved to be the same in the species analyzed. Only small differences were observed in the structure of the lateral and middle denticles.
|
|
| Figure 1. General morphology of digestive tract of Endecous spp. c = crop; es = esophagus; fg = foregut; g = gut; gc = gastric caecum; hg = hindgut; mg = midgut; Mt = Malpighian tube; ph = pharynx; pv = proventriculus; r = rectum; v = ventriculus. |
|
|
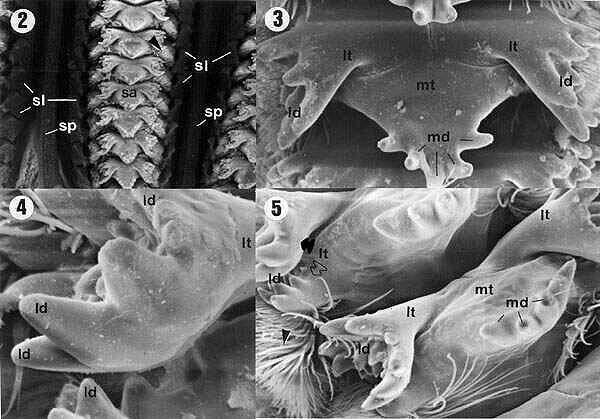
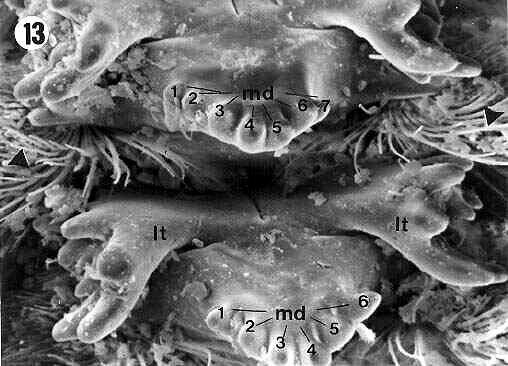
| Figures 2, 3, 4, 5. Internal view of the proventriculus of E. itatibensis from the cerrado region. ld = lateral denticles; lt = lateral teeth; md = middle denticles; mt = middle teeth; sa = sclerotized appendix; sl = sclerotized lobes; sp = sclerotized partitions; arrowheads = sclerotized setose lobes; filled arrow = external portion of lateral tooth, empty arrow = internal portion of a lateral tooth. Magnifications: 2 = 50X; 3 and 5 = 300X; 4 = 800X; 5 = 300X. |
2. General morphology of the proventriculus
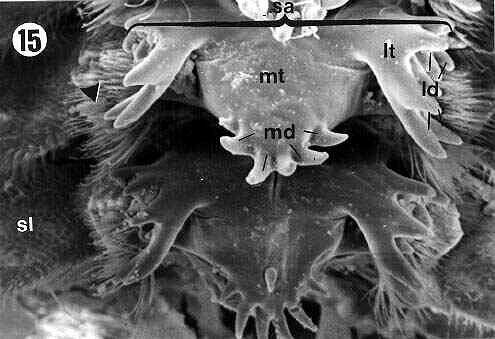
The inner surface of the proventriculus showed six rows of groups of sclerotized appendices (sa) connected to each other by sclerotized partitions (sp) (Figures 2, 9). On each side of the rows of sclerotized appendices (sa), small expansions were observed, the sclerotized lobes (sl) (Figures 2, 9, 15), which presented a dense internal setose (arrowheads in Figures 2, 5, 6, 7, 9, 13, 15).
The sclerotized appendix (sa) (Figures 2, 3, 6, 9, 13, 15) is formed by a central projection, the middle tooth (mt) (in detail on Figure 12), and two lateral expansions, the lateral teeth (lt) (details in the Figures 4, 8, 10, 11, 14, 16).
The middle tooth (mt) showed apical projections, the middle denticles (md) (Figures 3, 5, 6, 7, 12, 13, 15), which were presented in variable numbers depending on the specimens.
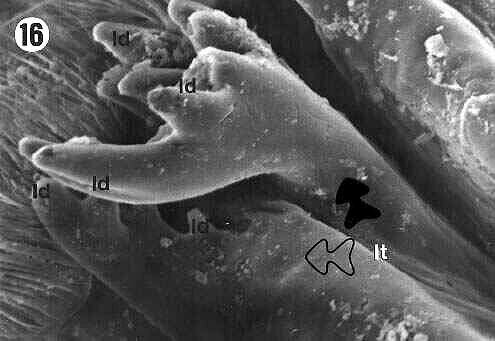
The lateral teeth (lt) are formed by two projections: a more external one, pointing backward to the lumen of the proventriculus (filled arrows in Figures 5, 10, 11, 14, 16) and a more internal one (empty arrows in Figures 5, 10, 11, 14, 16), both of them with apical projections called lateral denticles (ld) (details on Figures 4, 7, 8, 10, 11, 14, 16), whose extremities had a sharp or rounded shape.
3. Comparison of proventricular morphology
Although the general morphology of the proventriculus was the same for the three species analyzed, some small differences were observed among them in terms of number of middle denticles (md) and shape of the extremities of the middle (md) and lateral denticles (ld).
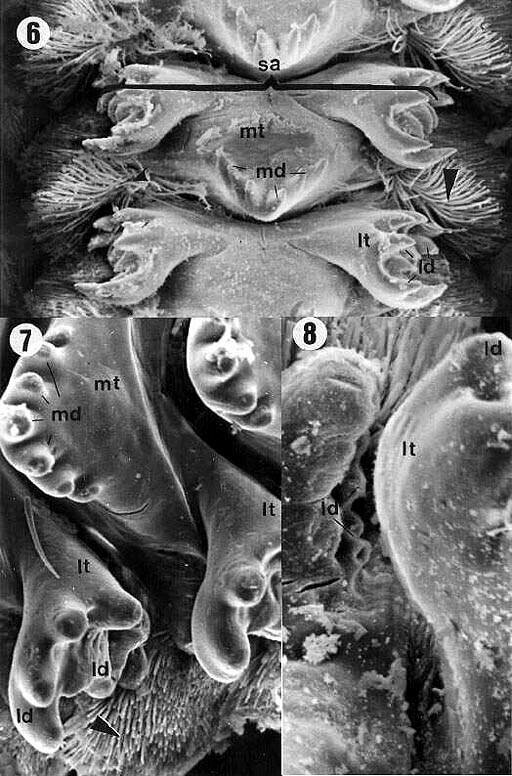
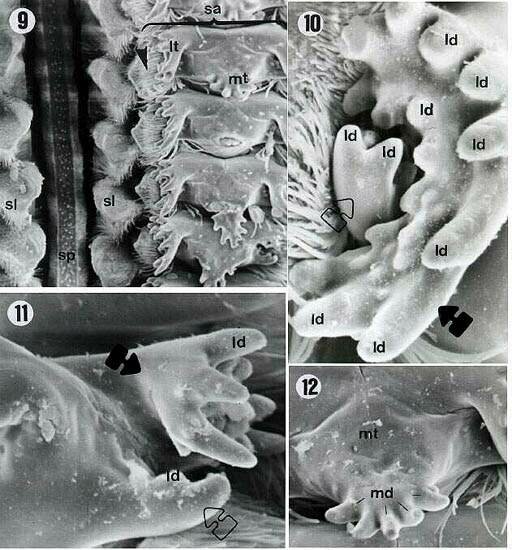
Six middle denticles (md) were observed in E.betariensis Figure 9). E. itatibensis showed five denticles in some specimens (figures 3, 5), six in others (Figures 6, 15), and an individual collected at Fazendão cave showed seven middle denticles (Figure 7). The species E. cavernicolus also presented variations in the number of middle denticles; some teeth with seven denticles and others with six were observed in the same individual (Figure 13).
The lateral (ld) and middle denticles (md) showed sharper extremities in all specimens of E. itatibensis from the cerrado (Figure 6) and from the forest of Itatiba (Figure 15); in all samples of E. itatibensis from the Fazendão cave (Figures 7, 8) and in the species E. betariensis (Figures 10, 12) and E. cavernicolus (Figure 13), the denticles were more rounded.
|
|
| Figures 6, 7, 8. Proventriculus of E. itatibensis from the cerrado region (figure 6) and Fazendão cave (figures 7, 8). ld = lateral denticles; lt = lateral teeth; md - middle denticles; mt = middle teeth; sa = sclerotized appendix; arrowheads = sclerotized setose lobes. Magnifications: 6 = 200X; 7 = 500X; 8 - 800X. |
|
|
| Figures 9, 10, 11, 12. E. betariensis proventriculus. ld = lateral denticles; lt = lateral tooth; md = middle teeth; sa = sclerotized appendix; sl = sclerotized lobes; sp = sclerotized partitions; empty arrow = internal portion of lateral teeth; filled arrows = external portion of the lateral teeth; arrowhead = sclerotized setose lobes. Magnifications: 9 = 200X; 10 = 1000X; 11 = 800X; 12 = 500X. |
|
|
| Figures 13, 14. E. cavernicolus proventriculus. ld = lateral denticles; lt = lateral teeth; md = middle denticles; filled arrow = external portion of the lateral teeth; empty arrow = internal portion of the lateral teeth; arrowheads = sclerotized setose lobes. Magnifications: 13 = 300X; 14 = 800X. |
|
|
| Figures 15, 16. E. itatibensis proventriculus from the Itatiba forest. sl = sclerotized lobe; sa = sclerotized appendix; lt = lateral teeth; mt = middle tooth; md = middle denticles; ld = lateral denticles; filled arrow = external portion of the lateral teeth; empty arrow = internal portion of the lateral teeth; arrowhead = sclerotized setose lobes. Magnifications: 15 = 300X; 16 = 800X. |
The great majority of crickets are omnivorous, but this does not necessarily imply that the diets of the different species are exactly the same. Within a genus, natural diets can be exclusively herbivorous or carnivorous, although few crickets are known as great predators, and when they are, they usually capture inactive or practically defenseless preys such as eggs, pupae, recently molted insects, etc. (Walker and Masaki 1989).
The natural diet of soil-dwelling crickets is not very well known; a few studies were reported; Gangwere (1961) observed dicotyledon leaves and other organic fragments in the content of the crop and pellets of Gryllus pennsylvanicus Burmeister, 1838 and Allonemobius allardi (Alexander and Thomas, 1958); he observed also smaller amounts of fragments of insects, pollen, spores and vegetables.
Bland and Rentz (1991) suggested that herbivorous gryllacridids have lateral denticles longer than those of predatory species; the authors point out that the high diversity of internal structures of the proventriculus among gryllacridid genera allows their use as taxonomic characters.
Species with a broad diet can have middle teeth able to grind, perforate and tear; this architecture combining different tooth and denticle shapes provides only general information but no detail about the feeding preferences of the species (Bland and Rentz 1991).
Isely (1944) and Gangwere (1965) observed diversity in the jaw and maxillary morphology of grasshoppers and related these differences to diet. Rentz and John (1990) observed that the mandible and, to a lesser extent, the maxilla differ at the genus level in gryllacridids. Bland and Rentz (1991) suggested that herbivorous species show a predominance of long lateral denticles in proventriculus when compared with predator species; the same was observed by Fontanetti and Zefa (2000) in Gryllus assimilis Fabricius, 1775.
The diet of Endecous, as well as that of most crickets, is practically unknown. Cave-dwelling Endecous species are usually found in guano deposits, suggesting that this is one of the main foods of these insects. The forest and cerrado species present large amounts of sand grains and larger organic matter particles compared to the contents of the proventriculus of cave species.
The species analyzed here are probably omnivorous and show complex sclerotized appendices, with large middle and lateral teeth containing a large number of denticles. This structure provides the insect with an apparatus with multiple food-breaking functions.
Cave-dwelling species have denticles with more rounded tips compared with epigean species. This difference was also observed between E. itatibensis specimens from caves and from forest populations.
Although no studies are available on the digestive content of the species analyzed, it is clear that food available in the forest and in caves differs both in quality and texture. In forests, the food probably consists of more solid particles, while the guano found in the caves is a fluid food. The authors believe that the small difference observed in the proventriculus of Endecous analyzed in the present study, although subtle, may reflect an adaptation to the variations in diet to which these populations were submitted in epigean and hypogean regions. The difference could be from wear the structures, but all specimens of each population showed it.
We wish to thank Mônica Iamonte, Antonio T.Yabuki, Lucila de L.Segalla Franco and Wenilton Luis Daltro for technical assistance, and CNPq for financial support.
Bland, TG, Rentz DCF. 1991. Studies in Australian Gryllacrididae: the proventriculus as a taxonomic character. Invert Taxon 5:443-455.
Bordas L. 1896. Etude de l'armature masticatrice du gésier chez les Blattidae et les Gryllidae. Compt Rend Nat Acad Sci Fr 123:271-273.
Bordas L. 1898. L'appareil digestif des orthoptères et son importance pour la classification des orthoptères. Ann Sci Nat Zool 5:1-192.
Costa-Lima A. 1940. Um novo grilo cavernícola de Minas Gerais, Orthoptera: Grylloidea: Phalangopsidae. Pap Avuls Depto Zool, Secretaria da Agric. S. Paulo, Brasil 1:43-50.
Deligne J. 1971. Morphologie du tube digestif et affinités systématiques du genre Tuberculitermes. Rev Zool Bot Afr 84(1-2):184-191.
Fontanetti CS, Zefa E. 2000. Morphological characterization of the proventriculus of Gryllus assimilis Fabricius (Orthoptera, Gryllidae).Rev Bras Zool 17(1):193-198.
Gangwere SK. 1961. A monograph on food selection in Orthoptera. Trans Am Entomol Soc 87:67-230.
Gangwere SK. 1965. The structural adaptations of mouthparts in Orthoptera and allies. EOS-Rev Esp Entomol 41:67-85.
Gibbs DG. 1967. The proventriculus of some trichopterous larvae. J Zool Lond 152:245-256.
Grant Jr HJ, Rentz DC. 1967. A biosystematic review of the family Tanaoceridae including a comparative study of the proventriculus. Pan-Pac Entomol 43(1):65-74.
Isely FB. 1944. Correlation between mandibular morphology and food specificity in grasshoppers. Ann Entomol Soc Am 37:47-67.
Judd WW. 1948. A comparative study of the proventriculus of orthopteroid insects with references to its use in taxonomy. Can J Res 26(D):93-161.
Karnowsky MJ. 1965. A formaldehyde-glutaraldehyde fixative at high osmolarity for use in electron microscopy. J Cell Biol 11:137-140.
Lebrun D. 1985. Structure digestives et régimes alimentaires des termites. Actes Coll Ins Soc 2:43-44.
Lebrun D, Lequet A. 1985. Relations entre les régimes alimentaires et la structure du gésier des termites. Bull Soc Sc Nat Fr 7(3):126-139.
Mello FAG, Pellegatti-Franco F. 1998. A new cave cricket of the genus Endecous from southeastern Brazil and characterization of male and female genitalia of E. itatibensis Rehn, 1918 (Orthoptera, Grylloidea, Phalangopsinae, Luzarinae). J Orth Res 7:185-188.
Mortiner TJ. 1965. The alimentary canal of some adult Lepidoptera and Trichoptera. Trans R Entomol Soc Lond 117(3):67-93.
Otte D, Alexander RD. 1983. The Australian crickets Orthoptera: Gryllidae. Lawrence: Allen Press, 477p. (Academy of Natural Sciences of Philadelphia, Monography 22).
Rehn JAG. 1918. On Dermatoptera and Orthoptera from South Eastern Brazil. Trans Am Entomol Soc 40:181-182.
Rentz DCF, John B. 1990. Studies in Australian Gryllacrididae: taxonomy, biology, ecology and cytology. Invert Taxon 3:1053-210.
Singh SB, Judd WW. 1966. A comparative study of the alimentary canal of adult calyptrate Diptera. Proc Entomol Soc Ontario 96:29-80.
Walker TJ, Masaki S. 1989. Natural History. In: Hubber F,; Moore TE, Lower W, editors. Cricket Behavior and Neurobiology. London: Cornell University Press. p. 1-42.